
Glutamate and glutamate receptors in the vertebrate retina by Victoria Connaughton – Webvision
Published by Reblogs - Credits in Posts,
The Organization of the Retina and Visual System
Glutamate and glutamate receptors in the vertebrate retina by Victoria Connaughton
Victoria Connaughton
1. General overview of synaptic transmission.
Cells communicate with each other electrically, through gap junctions, and chemically, using neurotransmitters. Chemical synaptic transmission allows nerve signals to be exchanged between cells which are electrically isolated from each other. The chemical messenger, or neurotransmitter, provides a way to send the signal across the extracellular space, from the presynaptic neuron to the postsynaptic cell. The space is called a cleft and is typically more than 10 nanometers across. Neurotransmitters are synthesized in the presynaptic cell and stored in vesicles in presynaptic processes, such as the axon terminal. When the presynaptic neuron is stimulated, calcium channels open and the influx of calcium ions into the axon terminal triggers a cascade of events leading to the release of neurotransmitter. Once released, the neurotransmitter diffuses across the cleft and binds to receptors on the postsynaptic cell, allowing the signal to propagate. Neurotransmitter molecules can also bind onto presynaptic autoreceptors and transporters, regulating subsequent release and clearing excess neurotransmitter from the cleft. Compounds classified as neurotransmitters have several characteristics in common (reviewed in Massey, 1990, Erulkar, 1994). Briefly, (1) the neurotransmitter is synthesized, stored, and released from the presynaptic terminal. (2) Specific neurotransmitter receptors are localized on the postsynaptic cells, and (3) there exists a mechanism to stop neurotransmitter release and clear molecules from the cleft. Common neurotransmitters in the retina are glutamate, GABA, glycine, dopamine, and acetylcholine. Neurotransmitter compounds can be small molecules, such as glutamate and glycine, or large peptides, such as vasoactive intestinal peptide (VIP). Some neuroactive compounds are amino acids, which also have metabolic functions in the presynaptic cell.
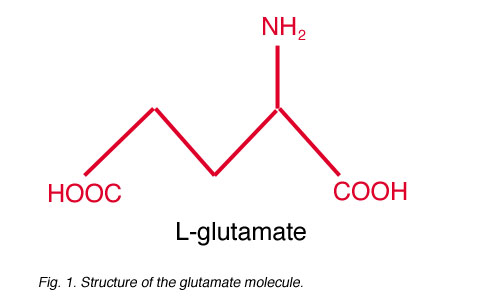
Fig. 1. Structure of the glutamate molecule
Glutamate (Fig. 1) is believed to be the major excitatory neurotransmitter in the retina. In general, glutamate is synthesized from ammonium and alpha-ketoglutarate (a component of the Krebs Cycle) and is used in the synthesis of proteins, other amino acids, and even other neurotransmitters (such as GABA; Stryer, 1988). Though glutamate is present in all neurons, only a few are glutamatergic, releasing glutamate as their neurotransmitter. Neuroactive glutamate is stored in synaptic vesicles in presynaptic axon terminals (Fykse and Fonnum, 1996). Glutamate is incorporated into the vesicles by a glutamate transporter located in the vesicular membrane. This transporter selectively accumulates glutamate through a sodium-independent, ATP-dependent process (Naito and Ueda, 1983, Tabb and Ueda, 1991, Fykse and Fonnum, 1996), resulting in a high concentration of glutamate in each vesicle. Neuroactive glutamate is classified as an excitatory amino acid (EAA) because glutamate binding onto postsynaptic receptors typically stimulates, or depolarizes, the postsynaptic cells.
2. Histological techniques identify glutamatergic neurons.

Fig. 2. Glutamate immunoreactivity
Using immunocytochemical techniques, neurons containing glutamate are identified and labeled with a glutamate antibody. In the retina, photoreceptors, bipolar cells, and ganglion cells are glutamate immunoreactive (Ehinger et al, 1988, Marc et al., 1990, Van Haesendonck and Missotten, 1990, Kalloniatis and Fletcher, 1993, Yang and Yazulla, 1994, Jojich and Pourcho, 1996) (Fig. 2). Some horizontal and/or amacrine cells can also display weak labeling with glutamate antibodies (Ehinger et al., 1988, Marc et al., 1990, Jojich and Pourcho, 1996; Yang, 1996). These neurons are believed to release GABA, not glutamate, as their neurotransmitter (Yazulla, 1986), suggesting the weak glutamate labeling reflects the pool of metabolic glutamate used in the synthesis of GABA. This has been supported by the results from double-labeling studies using antibodies to both GABA and glutamate: glutamate-positive amacrine cells also label with the GABA antibodies (Jojich and Pourcho, 1996, Yang, 1996).
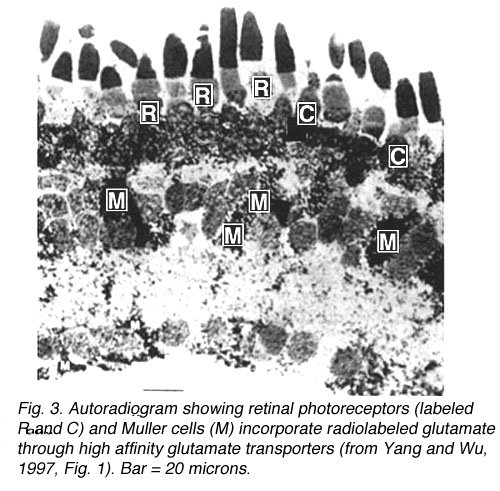
Fig. 3. Autoradiogram of glutamate uptake through glutamate transporters
Photoreceptors, which contain glutamate, actively take up radiolabeled glutamate from the extracellular space, as do Muller cells (Fig. 3) (Marc and Lam, 1981; Yang and Wu, 1997). Glutamate is incorporated into these cell types through a high affinity glutamate transporter located in the plasma membrane. Glutamate transporters maintain the concentration of glutamate within the synaptic cleft at low levels, preventing glutamate-induced cell death (Kanai et al., 1994). Though Muller cells take up glutamate, they do not label with glutamate antibodies (Jojich and Pourcho, 1996). Glutamate incorporated into Muller cells is rapidly broken down into glutamine, which is then exported from glial cells and incorporated into surrounding neurons (Pow and Crook, 1996). Neurons can then synthesize glutamate from glutamine (Hertz, 1979, Pow and Crook, 1996).
Thus, histological techniques are used to identify potential glutamatergic neurons by labeling neurons containing glutamate (through immunocytochemistry) and neurons that take up glutamate (through autoradiography). To determine if these cell types actually release glutamate as their neurotransmitter, however, the receptors on postsynaptic cells have to be examined.
3. Glutamate receptors.
Once released from the presynaptic terminal, glutamate diffuses across the cleft and binds onto receptors located on the dendrites of the postsynaptic cell(s). Multiple glutamate receptor types have been identified. Though glutamate will bind onto all glutamate receptors, each receptor is characterized by its sensitivity to specific glutamate analogues and by the features of the glutamate-elicited current. Glutamate receptor agonists and antagonists are structurally similar to glutamate (Fig. 4), which allows them to bind onto glutamate receptors. These compounds are highly specific and, even in intact tissue, can be used in very low concentrations because they are poor substrates for glutamate uptake systems (Tachibana and Kaneko, 1988, Schwartz and Tachibana, 1990).
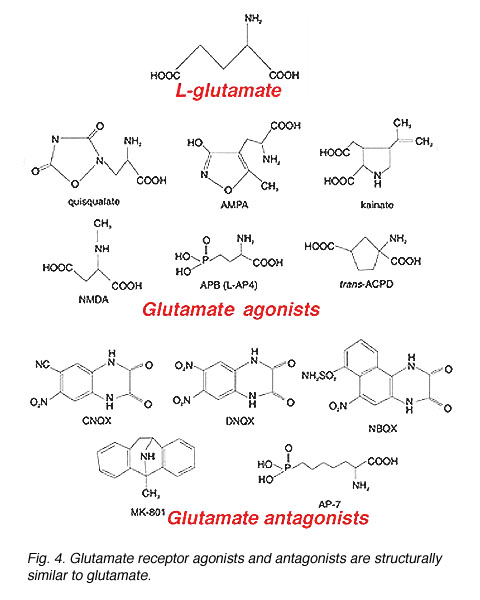
Fig. 4. Glutamate receptor agonists and antagonists
Two classes of glutamate receptors (Fig. 5) have been identified: (1) ionotropic glutamate receptors, which directly gate ion channels, and (2) metabotropic glutamate receptors, which may be coupled to an ion channel or other cellular functions via an intracellular second messenger cascade. These receptor types are similar in that they both bind glutamate and glutamate binding can influence the permeability of ion channels. However, there are several differences between the two classes.
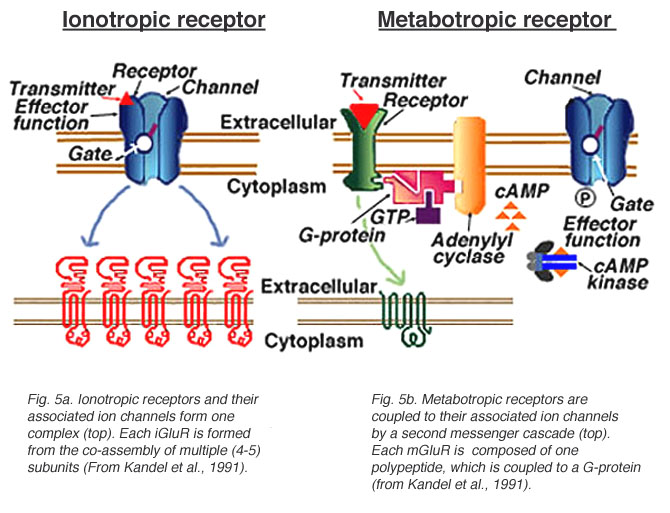
Fig. 5. Ionotropic and metabotropic glutamate receptors and channels
4. Ionotropic glutamate receptors.
Glutamate binding onto an ionotropic receptor directly influences ion channel activity because the receptor and the ion channel form one complex (Fig. 5a). These receptors mediate fast synaptic transmission between neurons. Each ionotropic glutamate receptor, or iGluR, is formed from the co-assembly of individual subunits. The assembled subunits may or may not be homologous, with the different combinations of subunits resulting in channels with different characteristics (Keinanen et al., 1990, Verdoorn et al., 1991, Moyner et al., 1992; Nakanishi, 1992, Ozawa and Rossier, 1996).
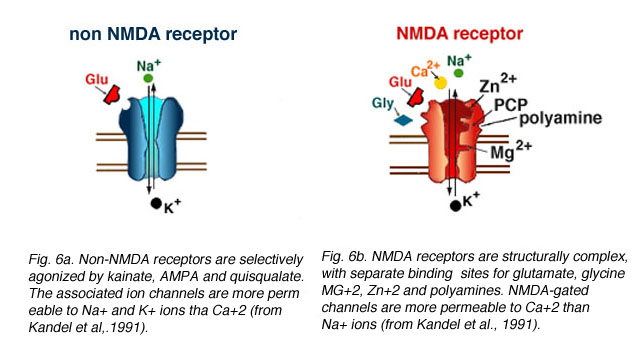
Fig. 6. Comparison between NMDA and non-NMDA receptors
Two iGluR types (see Fig. 6) have been identified: (1) NMDA receptors, which bind glutamate and the glutamate analogue N-Methyl-D-Aspartate (NMDA) and (2) non-NMDA receptors, which are selectively agonized by kainate, AMPA, and quisqualate, but not NMDA.
Non-NMDA receptors. Glutamate binding onto a non-NMDA receptor opens non-selective cation channels more permeable to sodium (Na+) and potassium (K+) ions than calcium (Ca+2) (Mayer and Westbrook, 1987). Glutamate binding elicits a rapidly activating inward current at membrane potentials negative to 0 mV, and an outward current at potentials positive to 0 mV. Kainate, quisqualate, and AMPA (alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid) are the specific agonists at these receptors; CNQX (6-cyano-7-nitroquinoxaline-2,3-dione), NBQX (1,2,3,4-tetrahydro-6-nitro-2,3-dione-benzo[f]quinoxaline-7-sulfonamide), and DNQX (6,7-dinitroquinoxaline-2,3-dione) are the antagonists.
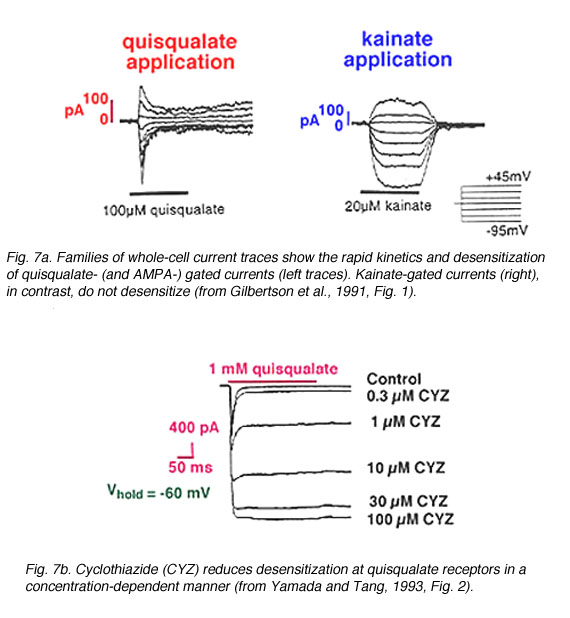
Fig. 7. Whole-cell patch clamp to show quisqualate and kainate gated currents
In retina, non-NMDA receptors have been identified on horizontal cells, OFF-bipolar cells, amacrine cells, and ganglion cells (see below). Patch clamp recordings (Gilbertson et al., 1991, Zhou et al., 1993, Boos et al., 1993, Cohen and Miller, 1994, Yu and Miller, 1995) indicate that AMPA, quisqualate, and/or kainate application can evoke currents in these cells. However, the kinetics of the ligand-gated currents differ. AMPA – and quisqualate-elicited currents rapidly desensitize; whereas, kainate-gated currents do not (Fig. 7a). The desensitization at AMPA/quisqualate receptors can be reduced (Fig. 7b) by adding cyclothiazide (Yamada and Tang, 1993), which stabilizes the receptor in an active (or non-desensitized) state (Yamada and Tang, 1993, Kessler et al., 1996).
Each non-NMDA receptor is formed from the co-assembly of several subunits (Boulter et al., 1990, Nakanishi et al., 1990, Nakanishi, 1992). To date, seven subunits (named GluR1 through GluR7) have been cloned (Hollmann et al., 1989, Boulter et al., 1990, Keinanen et al., 1990, Nakanishi et al., 1990, Bettler et al., 1990, 1992, Egebjerg et al., 1991). Expression of subunit clones in Xenopus oocytes revealed that GluR5, GluR6, and GluR7 (along with subunits KA1 and KA2) co-assemble to form kainate(-preferring) receptors; whereas, GluR1, GluR2, GluR3, and GluR4 are assembled into AMPA(-preferring) receptors (Nakanishi, 1992).
NMDA receptors. Glutamate binding onto an NMDA receptor also opens non-selective cation channels, resulting in a conductance increase. However, the high conductance channel associated with these receptors is more permeable to Ca+2 than Na+ ions (Mayer and Westbrook, 1987) and NMDA-gated currents typically have slower kinetics than kainate- and AMPA-gated channels. As the name suggests, NMDA is the selective agonist at these receptors. The compounds MK-801, AP-5 (2-amino-5-phosphonopentanoic acid), and AP-7 (2-amino-7-phosphoheptanoic acid) are NMDA receptor antagonists.
NMDA receptors are structurally complex, with separate binding sites for glutamate, glycine, magnesium ions (Mg+2), zinc ions (Zn+2), and a polyamine recognition site (Fig. 6b). There is also an antagonist binding site for PCP and MK-801 (Lodge, 1997). The glutamate, glycine, and magnesium binding sites are important for receptor activation and gating of the ion channel. In contrast, the zinc and polyamine sites are not needed for receptor activation, but affect the efficacy of the channel. Zinc blocks the channel in a voltage-independent manner (Westbrook and Mayer, 1987). The polyamine site (Ransom and Stec, 1988, Williams et al., 1994) binds compounds such as spermine or spermidine, either potentiating (Ranson and Stec, 1988; Williams et al., 1994) or inhibiting (Williams et al., 1994) the activity of the receptor, depending on the combination of subunits forming each NMDA receptor (Williams et al., 1994).
To date, five subunits (NR1, NR2a, N2b, N2c, and N2d) of NMDA receptors have been cloned (Moriyoshi et al., 1991, Ikeda et al., 1992, Katsuwada et al., 1992, Meguro et al., 1992, Ishii et al., 1993). As with non-NMDA receptors, NMDA receptor subunits can co-assemble as homomers (i.e., five NR1 subunits; Moyner et al., 1992, Moriyoshi et al., 1992) or heteromers (one NR1 + four NR2 subunits; Meguro et al., 1992, Katsuwada et al., 1992, Moyner et al., 1992, Ishii et al., 1993). However, all functional NMDA receptors express the NR1 subunit (Moyner et al., 1992, Nakanishi, 1992, Ishii et al., 1993).
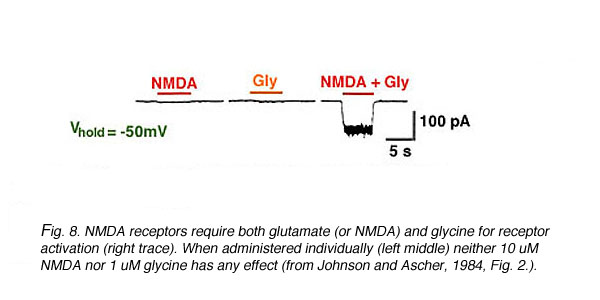
Fig. 8. NMDA receptor activation
The glutamate, glycine, and Mg+2 binding sites confer both ligand-gated and voltage-gated properties onto NMDA receptors. NMDA receptors are ligand-gated because the binding of glutamate (ligand) is required to activate the channel. In addition, micromolar concentrations of glycine must also be present (Fig. 8) (Johnson and Ascher, 1987, Kleckner and Dingledine, 1988). The requirement for both glutamate and glycine makes them co-agonists (Kleckner and Dingledine, 1988) at NMDA receptors.
Mg+2 ions provide a voltage-dependent block of NMDA-gated channels (Nowak et al., 1984). This can be seen in the current-voltage (I-V) relationship presented in Fig. 9 (from Nowak et al., 1984).
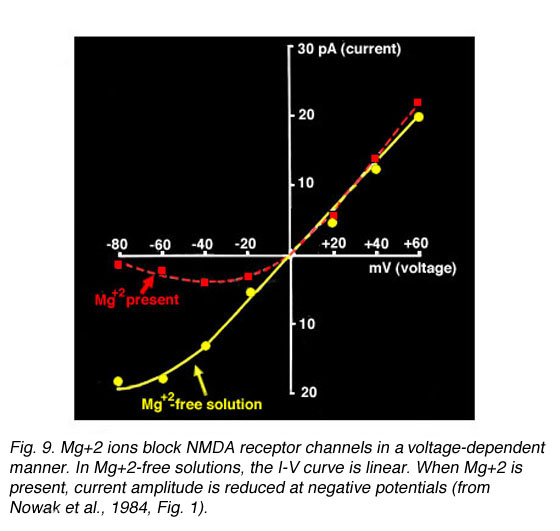
Fig. 9. Mg+2 ions block NMDA receptor channels
I-V curves plotted from currents recorded in the presence of Mg+2 have a characteristic J-shape (dotted line); whereas, a linear relationship is calculated in Mg+2-free solutions (solid line). At negative membrane potentials, Mg+2 ions occupy the binding site causing less current to flow through the channel. As the membrane depolarizes, the Mg+2 block is removed (Nowak et al., 1984).
Retinal ganglion cells and some amacrine cell types express functional NMDA receptors in addition to non-NMDA receptors (i.e., Massey and Miller, 1988, 1990, Mittman et al., 1990, Dixon and Copenhagen 1992, Diamond and Copenhagen, 1993, Cohen and Miller, 1994). The currents elicited through these different iGluR types can be distinguished pharmacologically. Non-NMDA receptor antagonists block a transient component of the ganglion cell light response; whereas, NMDA receptor antagonists block a more sustained component (Mittman et al., 1990, Diamond and Copenhagen, 1993, Hensley et al., 1993, Cohen and Miller, 1994). These findings suggest the currents elicited through co-localized NMDA and non-NMDA receptors mediate differential contributions to the ON- and OFF-light responses observed in ganglion cells (i.e., Diamond and Copenhagen, 1993).
5. Metabotropic glutamate receptors.
Unlike ionotropic receptors, which are directly linked to an ion channel, metabotropic receptors are coupled to their associated ion channel through a second messenger pathway. Ligand (glutamate) binding activates a G-protein and initiates an intracellular cascade (Nestler and Duman, 1994). Metabotropic glutamate receptors (mGluRs) are not co-assembled from multiple subunits, but are one polypeptide (Fig. 5b). To date, eight mGluRs (mGluR1-mGluR8) have been cloned (Houamed et al., 1991, Masu et al., 1991, Abe et al., 1992, Tanabe et al., 1992, Nakajima et al., 1993, Saugstad et al., 1994, Duvoisin et al., 1995). These receptors are classified into three groups (I, II, and III) based on structural homology, agonist selectivity, and their associated second messenger cascade (Table 1, end of chapter) (reviewed in Nakanishi, 1994, Knopfel et al., 1995, Pin and Duvoisin, 1995, Pin and Bockaert, 1995).
In brief, Group I mGluRs (mGluR1 and mGluR5) are coupled to the hydrolysis of fatty acids and the release of calcium from internal stores. Quisqualate and trans-ACPD are Group I agonists. Group II (mGluR2 and mGluR3) and Group III (mGluRs 4, 6, 7, and 8) receptors are considered inhibitory because they are coupled to the downregulation of cyclic nucleotide synthesis (Pin and Duvoisin, 1995). L-CCG-1 and trans-ACPD agonize Group II receptors; L-AP4 (also called APB) selectively agonizes Group III receptors. In situ hybridization studies have revealed that the mRNAs encoding Group I, II, and III mGluRs are present in retina (see below); however, with the exception of the APB receptor, the function of all these receptor types in retina has not been characterized.
APB receptor. In contrast to non-NMDA and NMDA receptors, glutamate binding onto an APB receptor elicits a conductance decrease (Slaughter and Miller, 1981, Nawy and Copenhagen, 1987, 1990) due to the closure of cGMP-gated non-selective cation channels (Nawy and Jahr, 1990) (Fig. 10).
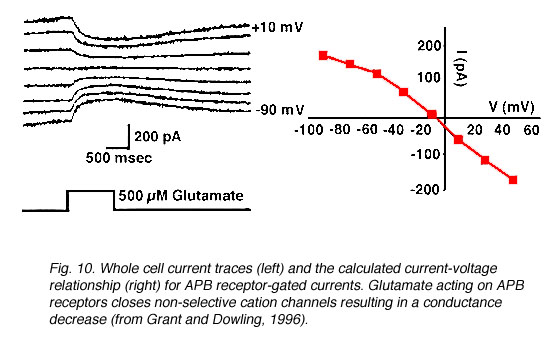
Fig. 10.Whole-cell current traces to show kinetics of APB receptor gated currents
APB application selectively blocks the ON-pathway in the retina (Fig. 11) (Slaughter and Miller, 1981), i.e., ON-bipolar cell responses and the ON-responses in amacrine cells (Taylor and Wassle, 1995) and ganglion cells (Cohen and Miller, 1994, Kittila and Massey, 1995, Jin and Brunken, 1996) are eliminated by APB. Experimental evidence (Slaughter and Miller 1981, Massey et al., 1983) suggests the APB receptor is localized to ON-bipolar cell dendrites. Inhibition of amacrine and ganglion cell light responses, therefore, is due to a decrease in the input from ON-bipolar cells, not a direct effect on postsynaptic receptors.

Fig. 11. Intracellular recordings to show that APB selectively antagonizes the ON-pathways
APB (2-amino-4-phosphobutyric acid, also called L-AP4) is the selective agonist for all Group III mGluRs (mGluR4, 6, 7, and 8). So, which is the APB receptor located on ON-bipolar cell dendrites? MGluR4, 7, and 8 expression has been observed in both the inner nuclear layer and the ganglion cell layer (Duvoisin et al., 1995, Hartveit et al., 1995) suggesting these mGluRs are associated with more than one cell type. In contrast, mGluR6 expression has been localized to the INL (Nakajima et al., 1993, Hartveit et al., 1995) and the OPL (Nomura et al., 1994) where bipolar cell somata and dendrites are located. Furthermore, ON-responses are abolished in mice lacking mGluR6 expression (Masu et al., 1995). These mutants also display abnormal ERG b-waves, suggesting an inhibition of the ON-retinal pathway at the level of bipolar cells (Masu et al., 1995). Taken together, these findings suggest the APB receptor on ON-bipolar cells is mGluR6.
6. Glutamate transporters and transporter-like receptors.
Glutamate transporters have been identified on photoreceptors (Marc and Lam, 1981, Tachibana and Kaneko, 1988, Eliasof and Werblin, 1993) and Muller cells (Marc and Lam, 1981, Yang and Wu, 1997). From glutamate labeling studies, the average concentration of glutamate in photoreceptors, bipolar cells, and ganglion cells is 5mM (Marc et al. 1990). Physiological studies using isolated cells indicate that only µM levels of glutamate are required to activate glutamate receptors (i.e., Aizenman et al., 1988, Zhou et al., 1993, Sasaki and Kaneko, 1996). Thus, the amount of glutamate released into the synaptic cleft is several orders of magnitude higher than the concentration required to activate most postsynaptic receptors. High affinity glutamate transporters located on adjacent neurons and surrounding glial cells rapidly remove glutamate from the synaptic cleft to prevent cell death (Kanai et al., 1994). Five glutamate transporters, EAAT-1 (or GLAST), EAAT-2 (or GLT-1), EAAT-3 (or EAAC-1), EAAT-4, and EAAT-5, have been cloned (Kanai and Hediger, 1992, Pines et al., 1992, Fairman et al., 1995, Schultz and Stell, 1996, Arriza et al., 1997, Kanai et al., 1997).
Glutamate transporters are pharmacologically distinct from both iGluRs and mGluRs. L-glutamate, L-aspartate, and D-aspartate are substrates for the transporters (Brew and Attwell, 1987, Tachibana and Kaneko, 1988, Eliasof and Werblin, 1993); glutamate receptor agonists (Brew and Attwell, 1987, Tachibana and Kaneko, 1988, Schwartz and Tachibana, 1990, Eliasof and Werblin, 1993) and antagonists (Barbour et al., 1991, Eliasof and Werblin, 1993) are not. Glutamate uptake can be blocked by the transporter blockers dihydrokainate (DHKA) and DL-threo-beta-hydroxyaspartate (HA) (Barbour et al., 1991, Eliasof and Werblin 1993).
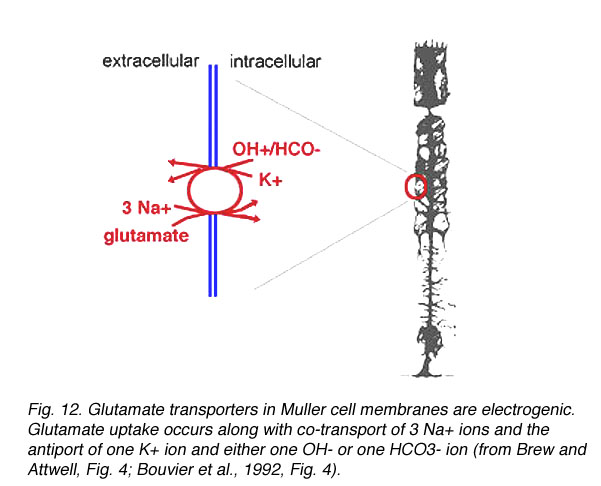
Fig. 12 Glutamate transporters in Muller cells are electrogenic
Glutamate transporters incorporate glutamate into Muller cells along with the co-transport of three Na+ ions (Brew and Attwell, 1987, Barbour et al., 1988) and the antiport of one K+ ion (Barbour et al., 1988, Bouvier et al., 1992) and either one OH- or one HCO3- ion (Bouvier et al., 1992) (Fig. 12). The excess sodium ions generate a net positive inward current which drives the transporter (Brew and Attwell, 1987, Barbour et al., 1988). More recent findings indicate a glutamate-elicited chloride current is also associated with some transporters (Eliasof and Jahr, 1996, Arriza et al., 1997).
It should be noted that the glutamate transporters located in the plasma membrane of neuronal and glial cells (discussed in this section) are different from the glutamate transporters located on synaptic vesicles within presynaptic terminals (see section 1). The transporters in the plasma membrane transport glutamate in a Na+- and voltage-dependent manner independent of chloride (Brew and Attwell, 1987, Barbour et al., 1988, Kanai et al., 1994). L-glutamate, L-aspartate, and D-aspartate are substrates for these transporters (i.e., Brew and Attwell, 1987). In contrast, the vesicular transporter selectively concentrates glutamate into synaptic vesicles in a Na+-independent, ATP-dependent manner (Naito and Ueda, 1983, Tabb and Ueda, 1991, Fykse and Fonnum, 1996) that requires chloride (Tabb and Ueda, 1991, Fykse and Fonnum, 1996).
Glutamate receptors with transporter-like pharmacology have been described in photoreceptors (Picaud et al., 1995a, b, Grant and Werblin, 1996) and ON-bipolar cells (Grant and Dowling 1995, 1996). These receptors are coupled to a chloride current. The pharmacology of these receptors is similar to that described for glutamate transporters, as the glutamate-elicited current is (1) dependent upon external Na+, (2) reduced by transporter blockers, and (3) insensitive to glutamate agonists and antagonists. However, altering internal Na+ concentration does not change the reversal potential (Picaud et al., 1995b) or the amplitude (Grant and Werblin, 1995, Grant and Dowling, 1996) of the glutamate-elicited current, suggesting the receptor is distinct from glutamate transporters. At the photoreceptor terminals, the glutamate-elicited chloride current may regulate membrane potential and subsequent voltage-gated channel activity (i.e., Picaud et al., 1995a). Postsynaptically, this receptor is believed to mediate conductance changes underlying photoreceptor input to ON- cone bipolar cells (Grant and Dowling, 1995).
7. Localization of glutamate receptor types in the retina.
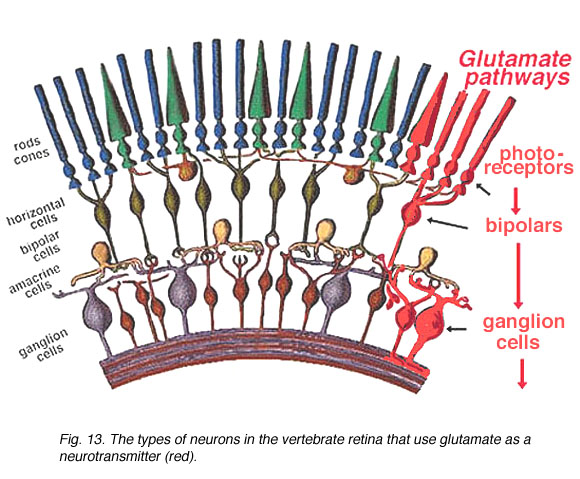
Fig. 13. The types of neurons in the vertebrate retina
Photoreceptor, bipolar, ganglion cells comprise the vertical transduction pathway in the retina. This pathway is modulated by lateral inputs from horizontal cells in the distal retina and amacrine cells in the proximal retina (Fig. 13). As described in the previous sections, photoreceptor, bipolar, and ganglion cells show glutamate immunoreactivity. Glutamate responses have been electrically characterized in horizontal and bipolar cells, which are postsynaptic to photoreceptors, and in amacrine and ganglion cells, which are postsynaptic to bipolar cells. Taken together, these results suggest glutamate is the neurotransmitter released by neurons in the vertical pathway. Recent in situ hybridization and immunocytochemical studies have localized the expression of iGluR subunits, mGluRs, and glutamate transporter proteins in the retina. These findings are summarized below.
8. Retinal neurons expressing ionotropic glutamate receptors.
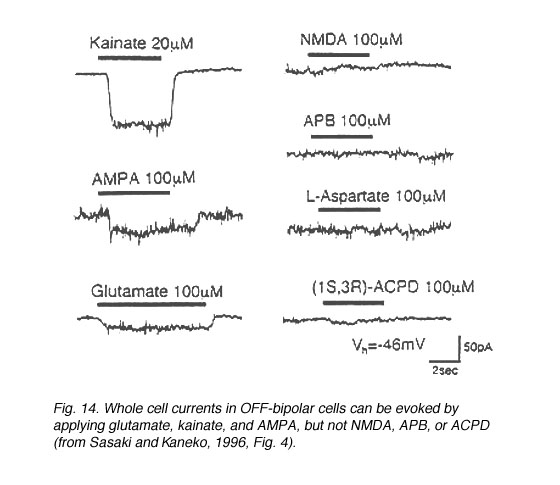 Fig.14. Whole-cell currents in OFF bipolar cells |
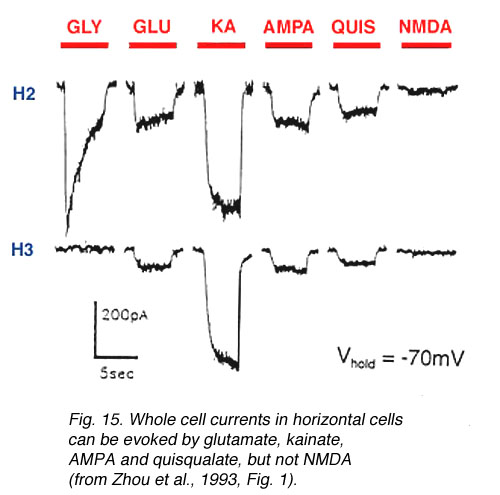 Fig. 15. Whole-cell currents in horizontal cells |
In both higher and lower vertebrates, electrophysiological recording techniques have identified ionotropic glutamate receptors on the neurons comprising the OFF-pathway (Table 2, end of chapter). In the distal retina, OFF-bipolar cells (Fig. 14) (Euler et al., 1996, Sasaki and Kaneko, 1996, Hartveit, 1997) and horizontal cells (Fig. 15) (Yang and Wu, 1991, Zhou et al., 1993, Kriaj et al., 1994) respond to kainate, AMPA, and quisqualate application, but not NMDA nor APB. (However, NMDA receptors have been identified on catfish horizontal cells (OÍDell and Christensen, 1989, Eliasof and Jahr, 1997) and APB-induced hyperpolarizations have been reported in some fish horizontal cells (Nawy et al., 1989, Takahashi and Copenhagen, 1992, Furukawa et al., 1997)).
Non-NMDA agonists also stimulate both amacrine cells (Fig. 16a) (Massey and Miller, 1988, Dixon and Copenhagen, 1992, Boos et al., 1993) and ganglion cells (Fig. 16b) (Mittman et al., 1990, Diamond and Copenhagen, 1993, Hensley et al., 1993, Cohen and Miller, 1994, Yu and Miller, 1995). Ganglion cells responses to NMDA have been observed (Massey and Miller, 1988, 1990, Mittman et al., 1990, Diamond and Copenhagen, 1993, Cohen and Miller, 1994); whereas, NMDA responses have been recorded in only some types of amacrine cells (Massey and Miller, 1988, Dixon and Copenhagen, 1992, Boos et al., 1993, but see Hartveit and Veruki, 1997).
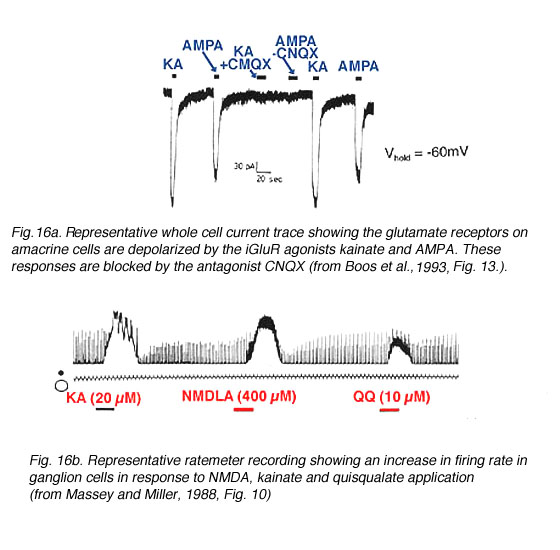
Fig. 16. Glutamate receptors on amacrine and ganglion cells
Consistent with this physiological data, antibodies to the different non-NMDA receptor subunits differentially label all retinal layers (Table 3, end of chapter; Hartveit et al., 1994, Peng et al., 1995, Hughes, 1997, Pourcho et al., 1997) and mRNAs encoding the different non-NMDA iGluR subunits are similarly expressed (Hughes et al., 1992, Hamassaki-Britto et al., 1993, Brandstatter et al., 1994). In contrast, mRNAs encoding NMDA subunits are expressed predominantly in the proximal retina, where amacrine and ganglion cells are located (INL, IPL, GCL; Table 3) (Brandstatter et al., 1994, Hartveit et al., 1994), though mRNA encoding the NR2a subunit (Hartveit et al., 1994) has been observed in the OPL and antibodies to the NR2d (Wenzel et al., 1997) and the NR1 subunits (Hughes, 1997) label rod bipolar cells.
9. Retinal neurons expressing metabotropic glutamate receptors.
All metabotropic glutamate receptors, except mGluR3, have been identified in retina either through antibody staining (Peng et al., 1995, Brandstatter et al., 1996, Koulen et al., 1997, Pourcho et al., 1997) or in situ hybridization (Nakajima et al., 1993, Duvoisin et al., 1995, Hartveit et al., 1995). MGluRs are differentially expressed throughout the retina, specifically in the outer plexiform layer, inner nuclear layer, inner plexiform layer, and the ganglion cell layer (Table 4, end of chapter). Though different patterns of mGluR expression have been observed in the retina, only the APB receptor on ON-bipolar cells has been physiologically examined.
10. Retinal neurons expressing glutamate transporters.
The glutamate transporters GLAST, EAAC1, and GLT-1have been identified in retina (Table 5, end of chapter). GLAST (L-glutamate/L-aspartate transporter) immunoreactivity is found in all retinal layers (Otori et al. 1994), but not in neuronal tissue. GLAST is localized to Muller cell membranes (Otori et al. 1994, Derouiche and Rauen, 1995, Rauen et al., 1996, Lehre et al., 1997). In contrast, EAAC-1 (excitatory amino acid carrier-1) antibodies do not label Muller cells or photoreceptors. EAAC-1 immunoreactivity is observed in ganglion and amacrine cells in chicken, rat, goldfish, and turtle retinas. In addition, bipolar cells positive labeled with EAAC-1 antibody in lower vertebrates and immunopositive horizontal cells were observed in rat (Schultz and Stell, 1996). GLT-1 (glutamate transporter-1) proteins have been identified in monkey (Grunert et al., 1994), rat (Rauen et al., 1996), and rabbit (Massey et al., 1997) bipolar cells. In addition, a few amacrine cells were weakly labeled with the GLT-1 antibody in rat (Rauen et al., 1996), as were photoreceptor terminals in rabbit (Massey et al., 1997).
11. Summary and conclusions.
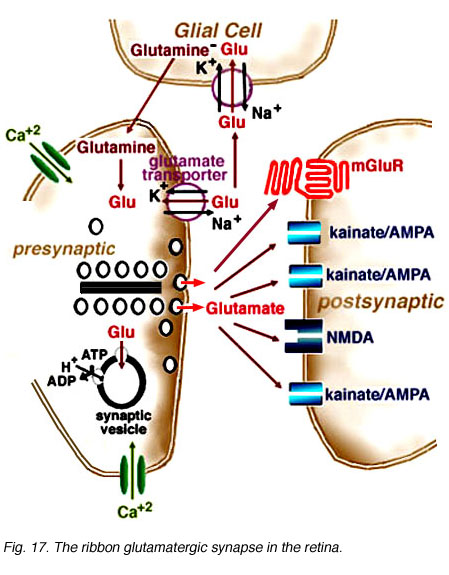
Fig. 17. The ribbon glutamatergic synapse in the retina
Histological analyses of presynaptic neurons and physiological recordings from postsynaptic cells suggest photoreceptor, bipolar, and ganglion cells release glutamate as their neurotransmitter. Multiple glutamate receptor types are present in the retina. These receptors are pharmacologically distinct and differentially distributed. IGluRs directly gate ion channels and mediate rapid synaptic transmission through either kainate/AMPA or NMDA receptors. Glutamate binding onto iGluRs opens cation channels, depolarizing the postsynaptic cell membrane. Neurons within the OFF-pathway (horizontal cells, OFF-bipolar cells, amacrine cells, and ganglion cells) express functional iGluRs. MGluRs are coupled to G-proteins. Glutamate binding onto mGluRs can have a variety of effects depending on the second messenger cascade to which the receptor is coupled. The APB receptor, found on ON-bipolar cell dendrites, is coupled to the synthesis of cGMP. At these receptors, glutamate decreases cGMP formation leading to the closure of ion channels. Glutamate transporters, found on glial and photoreceptor cells, are also present at glutamatergic synapses (Fig. 17). Transporters remove excess glutamate from the synaptic cleft to prevent neurotoxicity. Thus, postsynaptic responses to glutamate are determined by the distribution of receptors and transporters at a glutamatergic synapses which, in retina, determine the conductance mechanisms underlying visual information processing within the ON- and OFF-pathways.
Tables
Table 1
Metabotropic glutamate receptor groups (from Pin and Duvoisin, 1995).
| group | mGluR | agonist(s) | intracellular pathway | |
|---|---|---|---|---|
| I | mGluR1, mGluR5 | quisqualate, ACPD | increase phospholipase C activity, increase cAMP levels, increase protein kinase A activity | |
| II | mGluR2, mGluR3 | L-CCG-1, ACPD | decrease cAMP levels | |
| III | mGluR4, mGluR6. mGluR7, mGluR8 | L-AP4 (APB) | decrease cAMP or cGMP levels |
Table 2
Glutamate receptor types on retinal neurons, electrophysiological measurements.
| retinal cell type | non-NMDA receptor | NMDA receptor | mGluR | Glutamate receptor with transporter-like pharmacology | species | references |
|---|---|---|---|---|---|---|
| photoreceptors | ++ (cones) | salamander | Eliasof & Werblin, 1993; Picaud et al., 1995b | |||
| ++ (rods) | salamander | Grant & Werblin, 1996 | ||||
| OFF-bipolar cells | ++ | mudpuppy | Slaughter & Miller, 1981, 1983 | |||
| ++ | cat | Sasaki & Kaneko, 1996 | ||||
| ++ | salamander | Hensley et al., 1993 | ||||
| ++ | rat | Euler et al., 1996 | ||||
| ++ | mudpuppy | Slaughter & Miller, 1983 | ||||
| ON-bipolar cells | ++ | ++(APB) | mudpuppy | Slaughter & Miller, 1981, 1983 | ||
| ++(APB) | ++ | white perch | Grant & Dowling, 1995, 1996 | |||
| ++(APB) | salamander | Hirano & MacLeish, 1991 | ||||
| ++(L-AP4) | salamander | Hensley et al., 1993 | ||||
| ++(AP-4) | rat | Euler et al., 1996 | ||||
| ++(APB and cGMP) | salamander | Nawy & Jahr, 1990 | ||||
| ++(APB and cGMP) | cat | de la Villa et al., (1995) | ||||
| horizontal cells | ++ | white perch | Zhou et al., 1993 | |||
| ++ | mudpuppy | Slaughter & Miller, 1983 | ||||
| ++ | salamander | Yang & Wu, 1991 | ||||
| ++ | ++ | catfish | O’Dell & Christensen, 1989; Eliasof & Jahr, 1997 | |||
| amacrine cells | ++(AII) | rat | Boos et al., 1993 | |||
| ++ | ++ | mudpuppy | Slaughter & Miller, 1983 | |||
| ++ | ++ | rabbit | Massey & Miller, 1988 | |||
| ++ | ++ | rat | Hartveit & Veruki, 1997 | |||
| ++ (transient & sustained AC) | ++ (transient AC) | salamander | Dixon & Copenhagen, 1992 | |||
| ganglion cells | ++ | ++ | salamander | Diamond & Copenhagen, 1993; Mittman et al., 1990; Hensley et al., 1993 | ||
| ++ | ++ | primates | Cohen & Miller, 1994 | |||
| ++ | ++ | rat | Aizenman et al., 1988 | |||
| ++ | ++ | mudpuppy | Slaughter & Miller, 1983 | |||
| ++ | ++ | cat | Cohen et al., 1994 | |||
| ++ | ++ | rabbit | Massey & Miller, 1988; 1990 |
Table 3
Ionotropic glutamate receptor expression in retinal neurons and retinal layers, immunocytochemistry and in situ hybridization.
| retinal cell type or layer | non-NMDA receptor subunits | NMDA receptor subunits | species | references | |
|---|---|---|---|---|---|
| photoreceptors | GluR6/7 (single cone outer segments) | goldfish | Peng et al., 1995 | ||
| GluR1 (cone pedicles) | cat | Pourcho et al., 1997 | |||
| OPL | GluR2, GluR2/3, GluR6/7 | rat | Peng et al., 1995 | ||
| NR2A (punctate) | cat | Hartveit et al., 1994 | |||
| GluR2, GluR2/3 (photoreceptors) | goldfish | Peng et al., 1995 | |||
| bipolar cells | GluR2 (Mb cells) | goldfish | Peng et al., 1995 | ||
| GluR2, GluR2/3 | rat | Peng et al., 1995 | |||
| NR2D (RBC) | rat | Wenzel et al., 1997 | |||
| GluR2 and/or GluR4 | NR1 (RBC) | rat | Hughes, 1997 | ||
| GluR2 (RBC) | rat | Hughes et al., 1992 | |||
| horizontal cells | GluR6/7 | goldfish | Peng et al., 1995 | ||
| GluR2/3 | cat | Pourcho et al., 1997 | |||
| INL | GluR2/3, GluR6/7 | rat | Peng et al., 1995 | ||
| NR2A(inner) | rat | Hartveit et al., 1994 | |||
| GluR1, 2, 5 > GluR4 (outer third), GluR1, 2, 5 (middle third), GluR1-5 (inner third) | rat | Hughes et al. 1992 | |||
| GluR1-7 | rat, cat | Hamassaki-Britto et al., 1993 | |||
| KA2 (homogenous), GluR6 (inner), GluR7 (inner two thirds) | NR1 (homogenous), NR2A-B (inner third, patchy), NR2C (inner two-thirds) | rat | Brandstatter et al., 1994 | ||
| IPL | GluR1, GluR2/3, GluR6/7 | rat | Peng et al., 1995 | ||
| NR2A | rat, cat, rabbit, monkey | Hartveit et al., 1994 | |||
| amacrine cells | GluR6 | NR2A-C | rat | Brandstatter et al., 1994 | |
| GluR2/3 | cat | Pourcho et al., 1997 | |||
| GluR1, GluR2/3 | rat | Peng et al., 1995 | |||
| ganglion cells | GluR1 | rat | Peng et al., 1995 | ||
| GCL | GluR2/3, GluR6/7 | rat | Peng et al., 1995 | ||
| GluR1-5 | rat | Hughes et al., 1992 | |||
| GluR1-7 | rat, cat | Hamassaki-Britto et al., 1993 | |||
| GluR6-7, KA2 | NR1, NR2A-C | rat | Brandstatter et al., 1994 | ||
| Muller cells | GluR4 | rat | Peng et al., 1995 |
Table 4
Metabotropic glutamate receptor expression in retinal neurons and retinal layers, immunocytochemistry and in situ hybridization.
| retinal cell type or layer | Group 1 | Group II | Group III | species | references |
|---|---|---|---|---|---|
| OPL | mGluR1alpha, mGluR5a (RBC dendrites) | rat | Koulen et al., 1997 | ||
| mGluR6 (RBC dendrites) | rat | Nomura et al., 1994 | |||
| INL | mGluR8 | mouse | Duvoisin et al., 1995 | ||
| mGluR6 | rat | Nakajima et al., 1993 | |||
| mGluR5 (BC, HC), mGluR1 (AC) | mGluR2 (AC) | mGluR6 (RBC), mGluR7 (BC), mGluR4,7 (AC) | rat | Hartveit et al., 1995 | |
| IPL | mGluR1alpha | rat | Peng et al., 1995 | ||
| mGluR7 (CBC terminals; AC dendrites; few GC dendrites) | rat | Brandstatter et al., 1996 | |||
| mGluR1alpha, mGluR5a (AC dendrites) | rat | Koulen et al., 1997 | |||
| amacrine cells | mGluR1alpha | rat | Peng et al., 1995 | ||
| mGluR1alpha | mGluR2/3 | cat | Pourcho et al., 1997 | ||
| ganglion cells | mGluR1alpha | rat | Peng et al., 1995 | ||
| GCL | mGluR8 | mouse | Duvoisin et al., 1995 | ||
| mGluR1alpha | mGluR2/3 | cat | Pourcho et al., 1997 | ||
| mGluR1 | mGluR2 | mGluR4, 7 | rat | Hartveit et al., 1995 |
Table 5
Glutamate transporters in retinal neurons and retinal layers, immunocytochemical localizations.
| retinal cell type | EAAC-1 | GLAST | GLT-1 | species | reference |
|---|---|---|---|---|---|
| photoreceptors | + (cone soma to pedicles) | rabbit | Massey et al., 1997 | ||
| OPL | ++ | rat | Rauen et al., 1996 | ||
| ++ (rod spherules > cone pedicles) | rabbit | Massey et al., 1997 | |||
| horizontal cells | ++ | rat | Schultz & Stell, 1996; Rauen et al., 1996 | ||
| bipolar cells | ++ (2 types of CBCs) | rabbit | Massey et al., 1997 | ||
| ++ (faint) | ++ | rat | Rauen et al., 1996 | ||
| ++ | turtle, salamander | Schultz & Stell, 1996 | |||
| ++ (DB2, flat midget bipolar cells) | monkey | Grunert et al., 1994 | |||
| IPL | ++ (diffuse) | rabbit | Massey et al., 1997 | ||
| ++ | ++ | rat | Rauen et al., 1996 | ||
| ++ | goldfish, salamander, turtle, chicken, rat | Schultz & Stell, 1996 | |||
| amacrine cells | ++ | ++ | rat | Rauen et al., 1996 | |
| ++ | Schultz & Stell, 1996 | ||||
| ganglion cells | ++ | chicken, rat, goldfish, turtle | Schultz & Stell, 1996 | ||
| ++ | rat | Rauen et al., 1996 | |||
| Muller cells | ++ | rat | Rauen et al., 1996; Lehre et al., 1997; Deroiche & Rauen, 1995 |
References:
Abe T, Sugihara H, Nawa H, Shigemoto R, Mizuno N, Nakanishi S. Molecular characterization of a novel metabotropic glutamate receptor mGluR5 coupled to inositol phosphate/Ca2+ signal transduction. J Biol Chem. 1992;267:13361–13368. [PubMed]
Aizenman E, Frosch MP, Lipton SA. Responses mediated by excitatory amino acid receptors in solitary retinal ganglion cells from rat. J Physiol.1988;396:75–91. [PubMed] [Free Full text in PMC]
Arriza JL, Eliasof S, Kavanaugh MP, Amara SG. Excitatory amino acid transporter 5, a retinal glutamate transporter coupled to a chloride conductance.Proc Natl Acad Sci U S A. 1997;94:4155–4160. [PubMed] [Free Full text in PMC]
Barbour B, Brew H, Attwell D. Electrogenic glutamate uptake in glial cells is activated by intracellular potassium. Nature. 1988;335:433–435. [PubMed]
Barbour B, Brew H, Attwell D. Electrogenic uptake of glutamate and aspartate into glial cells isolated from the salamander (Ambystoma) retina. J Physiol.1991;436:169–193. [PubMed] [Free Full text in PMC]
Bettler B, Boulter J, Hermans-Borgmeyer I, O’Shea-Greenfield A, Deneris ES, Moll C, Borgmeyer U, Hollmann M, Heinemann S. Cloning of a novel glutamate receptor subunit, GluR5: expression in the nervous system during development. Neuron. 1990;5:583–595. [PubMed]
Bettler B, Egebjerg J, Sharma G, Pecht G, Hermans-Borgmeyer I, Moll C, Stevens CF, Heinemann S. Cloning of a putative glutamate receptor: a low affinity kainate-binding subunit. Neuron. 1992;8:257–265. [PubMed]
Boos R, Schneider H, Wassle H. Voltage- and transmitter-gated currents of AII amacrine cells in a slice preparation of the rat retina. J Neurosci.1993;13:2874–2888. [PubMed]
Boulter J, Hollmann M, O’Shea-Greenfield A, Hartley M, Deneris E, Maron C, Heinemann S. Molecular cloning and functional expression of glutamate receptor subunit genes. Science. 1990;249:1033–1037. [PubMed]
Bouvier M, Szatkowski M, Amato A, Attwell D. The glial cell glutamate uptake carrier countertransports pH-changing ions. Nature. 1992;360:471–474. [PubMed]
Brandstatter JH, Hartveit E, Sasso-Pognetto M, Wassle H. Expression of NMDA and high-affinity kainate receptor subunit mRNAs in the adult rat retina.Eur J Neurosci. 1994;6:1100–1112. [PubMed]
Brandstatter JH, Koulen P, Kuhn R, van der Putten H, Wassle H. Compartmental localization of a metabotropic glutamate receptor (mGluR7): two different active sites at a retinal synapse. J Neurosci. 1996;16:4749–4756. [PubMed]
Brew H, Attwell D. Electrogenic glutamate uptake is a major current carrier in the membrane of axolotl retinal glial cells. Nature. 1987;327:707–709. [PubMed]
Cohen ED, Miller RF. The role of NMDA and non-NMDA excitatory amino acid receptors in the functional organization of primate retinal ganglion cells. Vis Neurosci. 1994;11:317–332. [PubMed]
de la Villa P, Kurahashi T, Kaneko A. L-Glutamate-induced responses and cGMP-activated channels in three subtypes of retinal bipolar cells dissociated from the cat. J Neurosci. 1995;15:3571–3582. [PubMed]
Derouiche A, Rauen T. Coincidence of L-glutamate/L-aspartate transporter (GLAST) and glutamine synthetase (GS) immunoreactions in retinal glia: evidence for coupling of GLAST and GS in transmitter clearance. J Neurosci Res. 1995;42:131–143. [PubMed]
Diamond JS, Copenhagen DR. The contribution of NMDA and non-NMDA receptors to the light-evoked input-output characteristics of retinal ganglion cells. Neuron. 1993;11:725–738. [PubMed]
Dixon DB, Copenhagen DR. Two types of glutamate receptors differentially excite amacrine cells in the tiger salamander retina. J Physiol. 1992;449:589–606. [PubMed] [Free Full text in PMC]
Duvoisin RM, Zhang C, Ramonell K. A novel metabotropic glutamate receptor expressed in the retina and olfactory bulb. J Neurosci. 1995;15:3075–3083.[PubMed]
Egebjerg J, Bettler B, Hermans-Borgmeyer I, Heinemann S. Cloning of a cDNA for a glutamate receptor subunit activated by kainate but not AMPA.Nature. 1991;351:745–748. [PubMed]
Ehinger B, Ottersen OP, Storm-Mathisen J, Dowling JE. Bipolar cells in the turtle retina are strongly immunoreactive for glutamate. Proc Natl Acad Sci U S A. 1988;85:8321–8325. [PubMed] [Free Full text in PMC]
Eliasof S, Werblin F. Characterization of the glutamate transporter in retinal cones of the tiger salamander. J Neurosci. 1993;13:402–411. [PubMed]
Eliasof S, Jahr CE. Retinal glial cell glutamate transporter is coupled to an anionic conductance. Proc Natl Acad Sci U S A. 1996;93:4153–4158.[PubMed] [Free Full text in PMC]
Eliasof S, Jahr CE. Rapid AMPA receptor desensitization in catfish cone horizontal cells. Vis Neurosci. 1997;14:13–18. [PubMed]
Erulkar SD. Chemically mediated synaptic transmission: an overview. In: Siegel GJ, Agranoff BJ, Albers RW, Molinoff PB, editors. Basic neurochemistry, 5th ed. New York: Raven Press; 1994. p. 181-208.
Euler T, Schneider H, Wassle H. Glutamate responses of bipolar cells in a slice preparation of the rat retina. J Neurosci. 1996;16:2934–2944. [PubMed]
Fairman WA, Vandengerg RJ, Arriza JL, Kavanaugh MP, Amara SG. An excitatory amino-acid transporter with properties of a ligand-gated chloride channel. Nature. 1995;375:599–603. [PubMed]
Furukawa T, Yamada KM, Petruv R, Djamgoz MBA, Yasui S. Nitric oxide, 2-amino-4-phosphobutyric acid and light/dark adaptation modulate short-wavelength-sensitive synaptic transmission to retinal horizontal cells. Neurosci Res. 1997;27:65–74. [PubMed]
Fykse EM, Fonnum F. Amino acid neurotransmission: dynamics of vesicular uptake. Neurochem Res. 1996;21:1053–1060. [PubMed]
Gilbertson TA, Scobey R, Wilson M. Permeation of calcium ions through non-NMDA glutamate channels in retinal bipolar cells. Science. 1991;251:1613–1615. [PubMed]
Grant GB, Dowling JE. A glutamate-activated chloride current in cone-driven ON bipolar cells of the white perch retina. J Neurosci. 1995;15:3852–3862.[PubMed]
Grant GB, Werblin FS. A glutamate-elicited chloride current with transporter-like properties in rod photoreceptors of the tiger salamander. Vis Neurosci.1996;13:135–144. [PubMed]
Grant GB, Dowling JE. ON bipolar cell responses in the teleost retina are generated by two distinct mechanisms. J Neurophysiol. 1996;76:3842–3849.[PubMed]
Grunert U, Martin PR, Wassle H. Immunocytochemical analysis of bipolar cells in the macaque monkey retina. J Comp Neurol. 1994;348:607–627.[PubMed]
Hamassaki-Britto DE, Hermans-Borgmeyer I, Heinemann S, Hughes TE. Expression of glutamate receptor genes in the mammalian retina: the localization of GluR1 through GluR7 mRNAs. J Neurosci. 1993;13:1888–1898. [PubMed]
Hartveit E, Brandstatter JH, Sasso-Pognetto M, Laurie DJ, Seeburgh PH, Wassle H. Localization and developmental expression of the NMDA receptor subunit NR2A in the mammalian retina. J Comp Neurol. 1994;348:570–582. [PubMed]
Hartveit E, Brandsttter JH, Enz R, Wassle H. Expression of the mRNA of seven metabotropic glutamate receptors (mGluR1 to 7) in the rat retina. An in situ hybridization study on tissue section and isolated cells. Eur J Neurosci. 1995;7:1472–1483. [PubMed]
Hartveit E. Functional organization of cone bipolar cells in the rat retina. J Neurophysiol. 1997;77:1716–1730. [PubMed]
Hartveit E, Veruki ML. AII amacrine cells express functional NMDA receptors. Neuroreport. 1997;8:1219–1223. [PubMed]
Hensley SH, Yang X-L, Wu SM. Identification of glutamate receptor subtypes mediating inputs to bipolar cells and ganglion cells in the tiger salamander retina. J Neurophysiol. 1993;69:2099–2107. [PubMed]
Hertz L. Functional interactions between neurons and astrocytes. I. Turnover and metabolism of putative amino acid transmitters. Prog Neurobiol.1979;13:277–323. [PubMed]
Hirano AA, MacLeish PR. Glutamate and 2-amino-4-phosphobutyric acid evoke an increase in potassium conductance in retinal bipolar cells. Proc Natl Acad Sci U S A. 1991;88:805–809. [PubMed] [Free Full text in PMC]
Hollmann M, O’Shea-Greenfield A, Rogers SW, Heinemann S. Cloning by functional expression of a member of the glutamate receptor family. Nature.1989;342:643–648. [PubMed]
Houamed KM, Kuijper JL, Gilbert TL, Haldeman BA, O’Hara PJ, Mulvihill ER, Almers W, Hagen FS. Cloning, expression, and gene structure of a G protein-coupled glutamate receptor from rat brain. Science. 1991;252:1318–1321. [PubMed]
Hughes TE, Hermans-Borgmeyer I, Heinemann S. Differential expression of glutamate receptor genes (GluR1-5) in the rat retina. Vis Neurosci.1992;8:49–55. [PubMed]
Hughes TE. Are there ionotropic glutamate receptors on the rod bipolar cell of the mouse retina? Vis Neurosci. 1997;14:103–109. [PubMed]
Ikeda K, Nagasawa M, Mori H, Araki K, Sakimura K, Watanabe M, Inoue Y, Mishina M. Cloning and expression of the 4 subunit of the NMDA receptor channel. FEBS Lett. 1992;313:34–38. [PubMed]
Ishii T, Moriyoshi K, Sugihara H, Sakurada K, Kadotani H, Yokoi M, Akazawa C, Shigemoto R, Mizuno N, Masu M, Nakanishi S. Molecular characterization of the family of N-methyl-D-aspartate receptor subunits. J Biol Chem. 1993;268:2836–2843. [PubMed]
Jin XT, Brunken WJ. A differential effect of APB on ON- and OFF-center ganglion cells in the dark adapted rabbit retina. Brain Res. 1996;708:191–196.[PubMed]
Johnson JW, Ascher P. Glycine potentiates the NMDA response in cultured mouse brain neurons. Nature. 1987;325:529–531. [PubMed]
Jojich L, Pourcho RG. Glutamate immunoreactivity in the cat retina: a quantitative study. Vis Neurosci. 1996;13:117–133. [PubMed]
Kalloniatis M, Fletcher EL. Immunocytochemical localization of the amino acid neurotransmitters in the chicken retina. J Comp Neurol. 1993;336:174–193.[PubMed]
Kanai Y, Hediger MA. Primary structure and functional characterization of a high-affinity glutamate transporter. Nature. 1992;360:467–471. [PubMed]
Kanai Y, Smith CP, Hediger MA. A new family of neurotransmitter transporters: the high-affinity glutamate transporters. FASEB J. 1993;7:1450–1459.[PubMed]
Kanai Y, Trotti D, Nussberger S, Hediger MA. 1997. The high-affinity glutamate transporter family, structure, function, and physiological relevance. In: Reith MEA, editor. Neurotransmitter transporters: structure, function, and regulation. Totowa (NJ): Humana Press; 1997. p. 171-213.
Kandel ER, Schwartz JH, Jessell TM. . Principles of neuroscience. 3rd ed. New York: Elsevier Publishing Co. 1991
Katsuwada, T., Kashiwabuchi, N., Mori, H., Sakimura, K., Kushiya, E., Araki, K., Megure, H., Masaki, H., Kumanishi, T., Arakawa, M., and Mishina, M. (1992) Molecular diversity of the NMDA receptor channel. Nature, 358, 36-41. [PubMed]
Keinanen K, Wisden W, Sommer B, Werner P, Herb A, Versoorn TA, Sakmann B, Seeburg PH. A family of AMPA-selective glutamate receptors. Science.1990;249:556–560. [PubMed]
Kessler M, Arai A, Quan A, Lynch G. Effect of cyclothiazide on binding properties of AMPA-type glutamate receptors: lack of competition between cyclothiazide and GYKI 52466. Mol Pharmacol. 1996;49:123–131. [PubMed]
Kittila CA, Massey SC. Effect of ON pathway blockade on directional selectively in the rabbit retina. J Neurophysiol. 1995;73:703–712. [PubMed]
Kleckner NW, Dingledine R. Requirement for glycine activation of NMDA-receptors expressed in Xenopus oocytes. Science. 1988;241:835–837. [PubMed]
Knopfel T, Kuhn R, Allgeier H. Metabotropic glutamate receptors: novel targets for drug development. J Med Chem. 1995;38:1417–1426. [PubMed]
Koulen P, Kuhn R, Wassle H, Brandstatter JH. Group I metabotropic glutamate receptors mGluR1 and mGluR5a: localization in both synaptic layers of the rat retina. J Neurosci. 1997;17:2200–2211. [PubMed]
Krizaj D, Akopian A, Witkovsky P. The effects of L-glutamate, AMPA, quisqualate, and kainate on retinal horizontal cells depend on adaptational state: implications for rod-cone interactions. J Neurosci. 1994;14:5661–5671. [PubMed]
Lehre KP, Davanger S, Danbolt NC. Localization of the glutamate transporter protein GLAST in rat retina. Brain Res. 1997;744:129–137. [PubMed]
Lodge D. Subtypes of glutamate receptors. Historical perspectives on their pharmacological differentiation. In: Monaghan DT, Weinhold RJ, editors. The ionotropic glutamate receptors. Totowa (NJ): Humana Press; 1997. p. 1-38.
Marc RE, Lam DMK. Uptake of aspartic and glutamic acid by photoreceptors in goldfish retina. Proc Natl Acad Sci U S A. 1981;78:7185–7189. [PubMed] [Free Full text in PMC]
Marc RE, Liu W-LS, Kalloniatis M, Raiguel SF, Van Haesendonck E. Patterns of glutamate immunoreactivity in the goldfish retina. J Neurosci.1990;10:4006–4034. [PubMed]
Massey SC, Redburn DA, Crawford MLJ. The effects of 2-amino-4-phosphobutyric acid (APB) on the ERG and ganglion cell discharge of rabbit retina.Vision Res. 1983;23:1607–1613. [PubMed]
Massey SC, Miller RF. Glutamate receptors of ganglion cells in the rabbit retina: evidence for glutamate as a bipolar cell transmitter. J Physiol.1988;405:635–655. [PubMed] [Free Full text in PMC]
Massey SC. Cell types using glutamate as a neurotransmitter in the vertebrate retina. Prog Retinal Res. 1990;9:399–425.
Massey SC, Miller RF. N-Methyl-D-aspartate receptors of ganglion cells in rabbit retina. J Neurophysiol. 1990;63:16–30. [PubMed]
Massey SC, Koomen JM, Liu S, Lehre KP, Danbolt NC. Distribution of the glutamate transporter GLT-1 in the rabbit retina. Invest Ophthal Vis Sci.1997;38:S689.
Masu M, Tanabe Y, Tsuchida K, Shigemoto R, Nakanishi S. Sequence and expression of a metabotropic glutamate receptor. Nature. 1991;349:760–765.[PubMed]
Masu M, Iwakabe H, Tagawa Y, Miyoshi T, Yamashita M, Fukuda Y, Sasaki H, Hiroi K, Nakamura Y, Shigemoto R, Takada M, Nakamura K, Makao K, Katsuki M, Nakanishi S. Specific deficit of the ON response in visual transmission by targeted disruption of the mGluR6 gene. Cell. 1995;80:757–765.[PubMed]
Mayer ML, Westbrook GL. Permeation and block of N-methyl-D-aspartic acid receptor channels by divalent cations in mouse cultured central neurones. J Physiol. 1987;394:501–527. [PubMed] [Free Full text in PMC]
Meguro H, Mori H, Araki K, Kushiya E, Kutsuwada T, Yamazaki M, Kumanishi T, Arakawa M, Sakimura K, Mishina M. Functional characterization of a heteromeric NMDA receptor channel expressed from cloned cDNAs. Nature. 1992;357:70–74. [PubMed]
Mittman S, Taylor WR, Copenhagen DR. Concomitant activation of two types of glutamate receptor mediates excitation of salamander retinal ganglion cells. J Physiol. 1990;428:175–197. [PubMed] [Free Full text in PMC]
Moyner, H., Sprengel, R., Schoepfer, R., Herb, A., Higuchi, M., Lorneli, H., Burnashev, N., Sakmann, B., and Seeburg, P.H. (1992) Heteromeric NMDA receptors: molecular and functional distinction of subtypes. Science, 256, 1217-1221. [PubMed]
Moriyoshi K, Masu M, Ishii T, Shigemoto R, Mizuno N, Nakanishi S. Molecular cloning and characterization of the rat NMDA receptor. Nature.1991;354:31–37. [PubMed]
Naito S, Ueda T. Adenosine triphosphate-dependent uptake of glutamate into protein I-associated synaptic vesicles. J Biol Chem. 1983;258:696–699.[PubMed]
Nakajima Y, Iwakabe H, Akazawa C, Nawa H, Shigemoto R, Mizuno N, Nakanishi S. Molecular characterization of a novel retinal metabotropic glutamate receptor mGluR6 with a high agonist selectively for L-2-amino-4-phosphobutyrate. J Biol Chem. 1993;268:11868–11873. [PubMed]
Nakanishi N, Schneider NA, Axel R. A family of glutamate receptor genes: evidence for the formation of heteromultimeric receptors with distinct channel properties. Neuron. 1990;5:569–581. [PubMed]
Nakanishi S. Molecular diversity of glutamate receptors and implications for brain function. Science. 1992;258:597–603. [PubMed]
Nakanishi S. Metabotropic glutamate receptors: synaptic transmission, modulation, and plasticity. Neuron. 1994;13:1031–1037. [PubMed]
Nawy S, Copenhagen DR. Multiple classes of glutamate receptor on depolarizing bipolar cells in retina. Nature. 1987;325:56–58. [PubMed]
Nawy SE, Sie A, Copenhagen DR. The glutamate analog 2-amino-4-phosphonobutyrate antagonizes synaptic transmission from cones to horizontal cells in the goldfish retina. Proc Natl Acad Sci U S A. 1989;86:1726–1730. [PubMed] [Free Full text in PMC]
Nawy S, Copenhagen DR. Intracellular cesium separates two glutamate conductances in retinal bipolar cells of goldfish. Vision Res. 1990;30:967–972.[PubMed]
Nawy S, Jahr CE. Suppression by glutamate of cGMP-activated conductance in retinal bipolar cells. Nature. 1990;346:269–271. [PubMed]
Nestler EJ, Duman ES. G proteins and cyclic nucleotides in the nervous system. In: Siegel GJ, Agranoff BW, Albers RW, Molinoff PB, editors. Basic Neurochemistry. 5th ed. New York: Raven Press; 1994. p. 429-448.
Nomura A, Shigemoto R, Nakamura Y, Okamoto N, Mizuno N, Nakanishi S. Developmentally regulated postsynaptic localization of a metabotropic glutamate receptor in rat bipolar cells. Cell. 1994;77:361–369. [PubMed]
Nowak L, Bregestovski P, Ascher P, Herbet A, Prochiantz A. Magnesium gates glutamate-activated channels in mouse central neurones. Nature.1984;307:462–465. [PubMed]
O’Dell TJ, Christensen BN. Horizontal cells isolated from catfish retina contain two types of excitatory amino acid receptors. J Neurophysiol.1989;61:1097–1109. [PubMed]
Otori Y, Shimada S, Tanaka T, Ishimoto I, Tana Y, Tohyama M. Marked increase in glutamate-aspartate transporter (GLAST/GluT-1) mRNA following transient retinal ischemia. Brain Res Mol Brain Res. 1994;27:310–314. [PubMed]
Ozawa S, Rossier J. Molecular basis for functional differences of AMPA-subtype glutamate receptors. News Physiol Soc. 1996;11:77–82.
Peng YW, Blackstone CD, Huganir RL, Yau KW. Distribution of glutamate receptor subtypes in the vertebrate retina. Neuroscience. 1995;66:483–497.[PubMed]
Picaud S, Larsson HP, Wellis DP, Lecar H, Werblin F. Cone photoreceptors respond to their own glutamate release in the tiger salamander. Proc Natl Acad Sci U S A. 1995a;92:9417–9421. [PubMed] [Free Full text in PMC]
Picaud SA, Larsson HP, Grant GB, Lecar H, Werblin FS. Glutamate-gated chloride channel with glutamate-transporter-like properties in cone photoreceptors of the tiger salamander. J Neurophysiol. 1995b;74:1760–1771. [PubMed]
Pin JP, Bockaert J. Get receptive to metabotropic glutamate receptors. Curr Opin Neurobiol. 1995;5:342–349. [PubMed]
Pin JP, Duvoisin R. Review: Neurotransmitter receptors. I. The metabotropic glutamate receptors: structure and functions. Neuropharmacology.1995;34:1–26. [PubMed]
Pines G, Danbolt NC, Bjoras M, Zhang Y, Bendahan A, Eide L, Koepsell H, Storm-Mathisen J, Seeberg E, Kanner BI. Cloning and expression of a rat brain L-glutamate transporter. Nature. 1992;360:464–467. [PubMed]
Pourcho RG, Cai W, Qin P. Glutamate receptor subunits in cat retina: light and electron microscopic observations. Invest Ophthal Vis Sci. 1997;38:S46.
Pow DV, Crook DR. Direct immunocytochemical evidence for the transfer of glutamine from glial cells to neurons: use of specific antibodies directed against the D-stereoisomers of glutamate and glutamine. Neuroscience. 1996;70:295–302. [PubMed]
Ransom RW, Stec NL. Cooperative modulation of [3H}MK-801 binding to the N-methyl-D-aspartate receptor-ion channel complex by L-glutamate, glycine, and polyamines. J Neurochem. 1988;51:830–836. [PubMed]
Rauen T, Rothstein JF, Wassle H. Differential expression of three glutamate transporter subtypes in the rat retina. Cell Tissue Res. 1996;286:325–336.[PubMed]
Sasaki T, Kaneko A. L-Glutamate-induced responses in OFF-type bipolar cells of the cat retina. Vision Res. 1996;36:787–795. [PubMed]
Saugstad JA, Kinzie JM, Mulvihill ER, Segerson TP, Westbrook GL. Cloning and expression of a new member of the L-2-amino-4-phosphobutyric acid-sensitive class of metabotropic glutamate receptors. Mol Pharmacol. 1994;45:367–372. [PubMed]
Schultz K, Stell WK. Immunocytochemical localization of the high-affinity glutamate transporter, EAAC1, in the retina of representative vertebrate species.Neurosci Lett. 1996;211:191–194. [PubMed]
Schwartz EA, Tachibana M. Electrophysiology of glutamate and sodium co-transport in a glial cell of the salamander retina. J Physiol. 1990;426:43–80.[PubMed] [Free Full text in PMC]
Slaughter MM, Miller RF. 2-Amino-4-phosphobutyric acid: a new pharmacological tool for retina research. Science. 1981;211:182–184. [PubMed]
Slaughter MM, Miller RF. The role of excitatory amino acid transmitters in the mudpuppy retina: an analysis with kainic acid and N-methyl aspartate. J Neurosci. 1983;3:1701–1711. [PubMed]
Stryer L. . Biochemistry. 3rd ed. New York: W.H. Freeman. 1988
Tabb JS, Ueda T. Phylogenetic studies on the synaptic vesicle glutamate transporter. J Neurosci. 1991;11:1822–1828. [PubMed]
Tachibana M, Kaneko A. L-Glutamate-induced depolarization in solitary photoreceptors: a process that may contribute to the interaction between photoreceptors in situ. Proc Natl Acad Sci U S A. 1988;85:5315–5319. [PubMed] [Free Full text in PMC]
Takahashi K, Copenhagen DR. APB suppresses synaptic input to retinal horizontal cells in fish: a direct action on horizontal cells modulated by intracellular pH. J Neurophysiol. 1992;67:1633–1642. [PubMed]
Tanabe Y, Masu M, Ishii T, Shigemoto R, Nakanishi S. A family of metabotropic glutamate receptors. Neuron. 1992;8:169–179. [PubMed]
Taylor WR, Wassle H. Receptive field properties of starburst cholinergic amacrine cells in the rabbit retina. Eur J Neurosci. 1995;7:2308–2321. [PubMed]
Van Haesendonck E, Missotten L. Glutamate-like immunoreactivity in the retina of a marine teleost, the dragonet. Neurosci Lett. 1990;111:281–286.[PubMed]
Verdoorn TA, Burnashev N, Monyer H, Seeburg PH, Sakmann B. Structural determinants of ion flow through recombinant glutamate receptor channels.Science. 1991;252:1715–1718. [PubMed]
Wenzel A, Benke D, Mohler H, Fritschy J-M. N-Methyl-D-aspartate receptors containing the NR2D subunit in the retina are selectively expressed in rod bipolar cells. Neuroscience. 1997;78:1105–1112. [PubMed]
Westbrook GL, Mayer ML. Micromolar concentrations of Zn2+ antagonize NMDA and GABA responses of hippocampal neurons. Nature. 1987;328:640–643. [PubMed]
Williams K, Zappia AM, Pritchett DB, Shen YM, Molinoff PB. Sensitivity of the N-methyl-D-aspartate receptor to polyamines is controlled by NR2 subunits.Mol Pharmacol. 1994;45:803–809. [PubMed]
Yamada KA, Tang C-M. Benzothiadiazides inhibit rapid glutamate receptor desensitization and enhance glutamatergic synaptic currents. J Neurosci.1993;13:3904–3915. [PubMed]
Yang C-Y, Yazulla S. Glutamate-, GABA-, and GAD-immunoreactivities co-localize in bipolar cells of tiger salamander retina. Vis Neurosci. 1994;11:1193–1203. [PubMed]
Yang C-Y. Glutamate immunoreactivity in the tiger salamander retina differentiates between GABA-immunoreactive and glycine-immunoreactive amacrine cells. J Neurocytol. 1996;25:391–403. [PubMed]
Yang JH, Wu SM. Characterization of glutamate transporter function in the tiger salamander retina. Vision Res. 1997;37:827–838. [PubMed]
Yang X-L, Wu SM. Coexistence and function of glutamate receptor subtypes in the horizontal cells of the tiger salamander retina. Vis Neurosci.1991;7:377–382. [PubMed]
Yazulla S. GABAergic neurons in the retina. Prog Retinal Res. 1986;5:1–52.
Yu W, Miller RF. NBQX, an improved non-NMDA antagonist studied in retinal ganglion cells. Brain Res. 1995;692:190–194. [PubMed]
Zhou ZJ, Fain GL, Dowling JE. The excitatory and inhibitory amino acid receptors on horizontal cells isolated from the white perch retina. J Neurophysiol.1993;70:8–19. [PubMed]
Last updated May 7, 2007.
| The author Dr. Victoria Connaughton was born in Sellersville Pennsylvania. She received her B.A. from Bucknell University in Biology in 1989 and her Ph. D. in Marine Studies from The University of Delaware in 1994. She is currently an Assistant Professor in the Biology Department at American University, Washington D.C. In her thesis work under Dr. Charles Epifano, she studied the visually guided feeding behavior of larval fish. Dr. Connaughton pursued postdoctoral studies with Dr. Greg Maguire at the University of Houston, and Dr. Ralph Nelson at the National Institutes of Health. Dr. Connaughton’s current research interests include electrophysiological examination of zebrafish mutants with visual system defects and the characterization of light responses in zebrafish retinal bipolar cells. |

Search
Chapters
- Book
- Part I: Foundations
- Part II: Anatomy and Physiology of the retina
- The retinal pigment epithelium by Olaf Strauss
- Photoreceptors by Helga Kolb
- Outer Plexiform Layer by Helga Kolb
- Inner Plexiform Layer by Helga Kolb
- Morphology and Circuitry of Ganglion Cells by Helga Kolb
- Ganglion Cell Physiology by Ralph Nelson
- Melanopsin-expressing, Intrinsically Photosensitive Retinal Ganglion Cells (ipRGCs) by Dustin M. Graham and Kwoon Y. Wong
- Glial cells of the Retina by Helga Kolb
- The Architecture of the Human Fovea
- Part III: Retinal Circuits
- Circuitry for Rod Signals Through The Retina by Helga Kolb
- Cone Pathways through the Retina by Helga Kolb
- Roles of Amacrine Cells by Helga Kolb
- AII Amacrine Cells by Mahnoosh Farsaii and Victoria P. Connaughton
- Midget pathways of the primate retina underlie resolution and red green color opponency by Helga Kolb
- S-Cone Pathways by Helga Kolb
- Feedback Loops by Helga Kolb
- The Anatomy and Physiology of Direction-Selective Retinal Ganglion Cells by Jinyue Liu
- Myriad Roles for Gap Junctions in Retinal Circuits
- Part IV: Neurotransmitters in the Retina
- Part V: Phototransduction and Photoreceptor Synaptic Pathways
- Phototransduction in Rods and Cones by Yingbin Fu
- Glutamate and glutamate receptors in the vertebrate retina by Victoria Connaughton
- Bipolar Cell Pathways in the Vertebrate Retina by Ralph Nelson and Victoria Connaughton
- GABAc Receptors in the Vertebrate Retina by Haohua Qian
- S-Potentials and Horizontal Cells by Ido Perlman, Helga Kolb and Ralph Nelson
- Part VI: Retinal Neurogenesis: Early stages in the development of neurons and pathways
- Part VII: Color Vision
- Part VIII: Psychophysics of Vision
- Psychophysics of Vision by Michael Kalloniatis and Charles Luu
- Visual Acuity by Michael Kalloniatis and Charles Luu
- Temporal Resolution by Michael Kalloniatis and Charles Luu
- Light and Dark Adaptation by Michael Kalloniatis and Charles Luu
- Color Perception by Michael Kalloniatis and Charles Luu
- Space Perception by Michael Kalloniatis and Charles Luu
- Perception of Depth by Michael Kalloniatis and Charles Luu
- Part IX: Brain Visual Areas
- Part X: Repair and Regeneration in the visual sytem
- Part XI: Electrophysiology
- Part XII: Investigations of Human Retinal Disease
- Retinal Degeneration, Remodeling and Plasticity by Bryan William Jones, Robert E. Marc and Rebecca L. Pfeiffer
- Cellular Remodeling in Mammalian Retina Induced by Retinal Detachment by Steve Fisher, Geoffrey P. Lewis, Kenneth A Linberg, Edward Barawid and Mark V. Verardo
- Age-Related Macular Degeneration (AMD) by Gregory S. Hageman, Karen Gaehrs, Lincoln V. Johnson and Don Anderson
- Diabetic Retinopathy and A Novel Treatment Based On The Biophysics Of Rod Photoreceptors And Dark Adaptation by Geoffrey. B. Arden and David J. Ramsey
- What is glaucoma? by David Krizaj
- Part XIII: Facts and Figures concerning the human retina by Helga Kolb
- Part XIV: Evolution of Phototransduction, Vertebrate Photoreceptors and Retina by Trevor Lamb
- Part XV: Visual Prostheses
- Part XVI: Anterior Segment
- Part XVII: Refractive Errors
- Changelog
